Virologische Analysen in der Influenzasaison 2019/20
Die hier vorgestellten Ergebnisse des Nationalen Referenzzentrums für Influenzaviren (NRZI) umfassen Daten zu innerhalb des Sentinels der Arbeitsgemeinschaft Influenza (AGI) isolierten Viren und von Viren, die im Rahmen von Ausbrüchen, der Untersuchung schwerer Erkrankungsfälle und von Typisierungsanfragen oder Projekten untersucht wurden sowie von Isolaten aus Einsendungen von Instituten und Gesundheitsämtern. Untersuchungen zur weiterführenden Charakterisierung von Respiratorischen Synzytial-Viren (RSV) und weiteren respiratorischen Viren wurden darüber hinaus bei einer Subpopulation der Sentinelproben vom Konsiliarlabor (KL) für RSV, Parainfluenzaviren (PIV) und Humane Metapneumoviren (HMPV) durchgeführt. Ab der 9. Kalenderwoche 2020 wurden Untersuchungen auf SARS-CoV-2 integriert.
In der Saison 2019/20 (KW 40/2019-KW20/2020) und der nachfolgenden Sommersurveillance (bis KW 39/2020) wurden im NRZI insgesamt 4637 Sentinelproben untersucht. In 2290 Proben wurden respiratorische Viren mittels real-time PCR (qPCR) nachgewiesen. Unter den 919 (40% der respiratorischen Viren) nachgewiesenen Influenzaviren waren 375 (41%) A(H1N1)pdm09- und 415 (45%) A(H3N2)- Viren sowie 127 (14%) B-Influenzaviren der Victoria-Linie und ein B-Influenzavirus der Yamagata-Linie sowie ein zoonotisches A/sw/H1avN1-Influenzavirus aus einer Übertragung von Schweinen. Zusätzlich wurden 834 (36% der respiratorischen Viren) Rhinoviren, 201 (9%) RSV, 243 (11%) HMPV, PIV1, 2, 3 und 4 sowie 14 (0,6%) SARS-CoV-2 nachgewiesen. In einigen Proben wurden Mehrfachinfektionen detektiert.
Weitere 119 Proben wurden im Projekt Surveillance schwerer akuter respiratorischer Infektionen untersucht (SARI-Surveillance), 190 Proben im Projekt RSV sowie 104 Proben aus Auftragsuntersuchungen. 472 Influenzavirusisolate wurden von Laboratorien zur weiteren Charakterisierung an das NRZI eingesandt.
Eine Auswahl repräsentativer Influenzaviren wurde während der Saison zum Referenzlabor der Weltgesundheitsorganisation (WHO) nach London gesandt für vergleichende Untersuchungen im Rahmen der Mitwirkung an der weltweiten virologischen Influenzavirus-Surveillance.

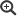 Abb. 1: Anzahl untersuchter Proben und Nachweisraten respiratorischer Viren in Proben des Sentinels der Arbeitsgemeinschaft Influenza in der Saison 2019/20
Quelle: RKI
Abb. 1: Anzahl untersuchter Proben und Nachweisraten respiratorischer Viren in Proben des Sentinels der Arbeitsgemeinschaft Influenza in der Saison 2019/20
Quelle: RKI
Die Saison 2019/20 in Deutschland war durch eine parallele dominante Zirkulation von A(H1N1)pdm09- und A(H3N2)-Influenzaviren gekennzeichnet, die durch eine geringgradige Zirkulation von Influenza-B-Viren der Viktoria-Linie begleitet wurde. Mit der Einführung von Schutzmaßnahmen zur Prävention der Ausbreitung von SARS-CoV-2 nahmen die Influenzavirusnachweise und die anderer respiratorischer Viren rapide ab (Abb. 1). Ab KW 15 wurden keine Influenzaviren mehr nachgewiesen. Lediglich im Juni kam es zum Nachweis eines zoonotischen Virus und im September zu zwei Nachweisen von saisonalen Influenzaviren.
Anzucht und antigene Charakterisierung
In der Saison 2019/20 wurden 572 Influenzaviren in Zellkultur isoliert, darunter 246 (43%) A(H3N2)-, 234 (41%) A(H1N1)pdm09-, 89 (16%) B/Victoria-Influenzaviren und ein B/Yamagata-Influenzavirus sowie ein H1N1-Influenzavirus aus einer zoonotischen Übertragung nach Exposition zu Schweinen. Weitere Influenzavirusisolate wurden von Instituten und Laboren zur Verfügung gestellt.
413 A(H1N1)pdm09-, 340 A(H3N2)-, 125 B/Victoria- und zwei B/Yamagata-Influenzaviren wurden mit Hilfe spezifischer Immunseren (Frettchen) im Hämagglutinationshemmtest (HHT) hinsichtlich ihres antigenen Profils charakterisiert. Der HHT wurde mit Putenerythrozyten durchgeführt. Für die Etablierung der Immunseren wurden Impfstämme und Referenzviren der WHO verwendet (Impfstämme der Saison 2019/20).
Die Untersuchungen dienen der Prüfung der Passfähigkeit der von der WHO empfohlenen Impfstoffe. Sie ermöglichen keine Aussagen zur Wirksamkeit der Impfstoffe, da für diese weitere Aspekte wie Antigengehalt in der Impfdosis, Impfschema, Adjuvans, die durch den jeweiligen Impfstamm induzierte Dauer der Immunität und Status des Impflings (Alter, vorhergehende Antigenkontakte zu Influenzaviren, immunologische Reaktivität) von Bedeutung sind.
In der Saison 2019/20 reagierten alle untersuchten Influenzavirusisolate mit den entsprechenden Immunseren, jedoch gab es Abweichungen in Bezug auf die Passgenauigkeit (Abb. 2). Diese waren besonders deutlich bei A(H1N1)pdm09-Influenzaviren, von denen 1/3 um mehr als 2 log2 schwächer reagierten als die homologen Impfstammantiseren. Aufgrund der starken Immunogenität der A(H1N1)pdm09-Influenzaviren bei Menschen hatte dies keine drastischen Auswirkungen, weil diese Viren mit reduzierter Reaktivität immer noch im Bereich der Reaktivität der anderen, schwächer antigen reagierenden Influenzaviren lagen. Keine Abweichungen der Passgenauigkeit gab es bei A(H3N2)- und B/Yamagata-Influenzaviren und wenige bei B/Victoria-Influenzaviren. Eine verringerte Passgenauigkeit der Influenzaviren ist häufig auf Mutationen in den antigenen Bindungsstellen zurückzuführen. Viren mit Mutationen in den antigenen Bindungsstellen werden durch das Immunsystem geimpfter Personen nicht so gut erkannt, was sich auf die Dauer der Immunität der Impfung auswirken kann. Die verringerte Passgenauigkeit der A(H1N1)pdm09-Influenzaviren ist auf die Mutation N156K in der Antigendomäne Sa des Hämagglutinins zurückzuführen, die der Influenzaviren der B-Victoria-Linie wird durch Aminosäureaustausche in den Positionen 128 und 133 (Antigendömane 120-loop) und in der Position 233 (nahe der Rezeptorbindungsdomäne 240-loop) begünstigt.
Insgesamt stimmen die Untersuchungen zur antigenen Charakterisierung der Influenzaviren der Saison 2019/20 gut mit den epidemiologischen Erhebungen zur Impfwirksamkeit überein, welche Punktschätzwerte für die klinische Wirksamkeit von 61% für A(H1N1)pdm09-, 62% für A(H3N2)- 73% für Influenza-B-Viren ergab1.
 Abb. 2: Passgenauigkeit der Impfstämme in der Influenzasaison 2019/20. Weitere Informationen zur Abb. 2 siehe: Erläuterung zur Passgenauigkeit der Impfstämme.
Quelle: RKI
Abb. 2: Passgenauigkeit der Impfstämme in der Influenzasaison 2019/20. Weitere Informationen zur Abb. 2 siehe: Erläuterung zur Passgenauigkeit der Impfstämme.
Quelle: RKI
Erläuterung zur Passgenauigkeit der Impfstämme (Abb. 2)
In der Abb. 2 sind die Ergebnisse der Passgenauigkeit der Impfstämme in der Influenzasaison 2019/20 dargestellt. Rhomben: isolierte Influenzaviren des jeweiligen Subtyps; dunkelblauer Balken: Reaktionsbreite des homologen Serums, hellblaue Klammer: optimale Passgenauigkeit (Toleranzbreite homologes Serum + tolerierte Abweichung von 2 log2). Bei allen Viren, die im Bereich der hellblauen Klammer liegen, ist von einer sehr hohen Passgenauigkeit auszugehen. Die eingesetzte Frettchen-Antiseren waren gegen folgende Impfviren gerichtet: A/Brisbane/2/2018 A(H1N1)pdm09, A/Kansas/14/2017 A(H3N2), B/Colorado/6/2017 der Victoria-Linie und B/Phuket/3073/13 der Yamagata-Linie.
nach oben
Molekulare Analysen
In der Saison 2019/20 wurde das Genom von 395 Influenzaviren sequenziert (überwiegend komplette Genome). Alle acht Segmente dieser Influenzaviren wurden phylogenetisch analysiert. Nachfolgend wird die Analyse des Hämagglutinin(HA)-Gens beschrieben, weil das Hämagglutinin das Hauptimmunogen ist. 164 (42%) A(H1N1)pdm09-Influenzaviren wurden sequenziert (156 AGI-Sentinel/6 Non-Sentinel/2 SARI-Surveillance), von denen 78 Viren phylogenetisch abgebildet wurden (HA: Nukleotidposition nt, Nummerierung vom ersten Nukleotid des Gens beginnend) nt84–nt1727). Von 169 (43%) sequenzierten A(H3N2)-Influenzaviren (155 AGI-Sentinel/5 Non-Sentinel/9 SARI-Surveillance) wurden 87 Influenzaviren phylogenetisch dargestellt (HA: nt78–nt1727). Da fast ausschließlich Influenza-B-Viren der Victoria-Linie zirkulierten, wurden nur diese Viren phylogenetisch untersucht (62 [16%] Viren: 57 AGI-Sentinel/2 Non-Sentinel/3 SARI-Surveillance, HA: nt79-nt1791). Die Anzahl repräsentiert dabei die unterschiedlich starke Zirkulation der Viren in der Saison 2019/20. Der überwiegende Anteil der charakterisierten Influenza-A- und -B-Viren stammte aus dem AGI-Sentinel. PCR-positive Sentinelproben wurden, bis auf wenige Ausnahmen, randomisiert für die Sequenzierung ausgewählt. Die molekulare Charakterisierung der Viren erfolgte über Next Generation Sequencing (NGS, Vollgenom-Analyse). Die identifizierten Substitutionen und Aminosäurepolymorphismen (ab 10% der minoritären Variante) wurden in den phylogenetischen Analysen erfasst. Die über NGS generierten Sequenzen sind unter www.gisaid.org hinterlegt.
Molekulare Analyse von Influenza A(H1N1)pdm09-Viren
In die Analysen wurden folgende A(H1N1)pdm09-Influenzaviren aus der aktuellen Saison aufgenommen und charakterisiert:
Ein großer Anteil aller Influenzanachweise war in der aktuellen Saison auf A(H1N1)pdm09-Influenzaviren zu¬rückzuführen (41% aller identifizierten Viren im AGI-Sentinel bis zur 13. KW 2020). Die phylogenetische Analyse der HA-Gene der Influenza-A(H1N1)pdm09-Viren zeigt, dass in Deutschland in der Saison 2019/20 überwiegend Viren der Gruppe 6B.1A5A nachgewiesen wurden. Auch auf internationaler Ebene wurden fast nur Viren der Gruppe 6B.1A5A (93% 2, 98% 3) detektiert, für die die Substitutionen HA1 S74R, S164T, I295V und überwiegend S183P (6B.1A), HA1 N260D (6B.1A5) sowie HA1 N129D, T185I (6B.1A5A, schwarz umkreist) charakteristisch sind 2, 3. Darüber hinaus wurden innerhalb der 6B.1A5A-Gruppe neue Varianten identifiziert, die fünf Subgruppen (grau umkreist) bilden mit den Aminosäuresubstitutionen HA1 D187A, Q189E (Subgruppe/SG 1); HA1 K130N, N156K, L161I, V250A, HA2 E179D (SG2); HA2 V193A (SG 3); HA1 N156K, A195E (SG 4) sowie HA1 R205K (SG 5). Weiterhin zirkulierten wenige 6B.1A7-Viren mit Aminosäureaustauschen in den Positionen HA1 K302T und HA2 I77M, N169S, E179D (blau umkreist), die zusätzlich im HA1 E68D, S121N, L161I sowie T120A aufwiesen (Abb. 3).
In der Gruppe aller analysierten A(H1N1)pdm09-Influenzaviren, die sich zu 95% aus dem AGI-Sentinel rekrutierten, wurden überwiegend die 6B.1A5A-Viren (98%; 161 von 164) sowie 6B.1A7-Viren (2%) nachgewiesen. Darüber hinaus wurden in der Saison 2019/20 bei den 6B.1A5A-Viren neue Varianten mit zusätzlichen Aminosäureaustauschen identifiziert, die zu unterschiedlichen Anteilen nachgewiesen wurden: HA1 D187A, Q189E (52%, SG 1), HA1 K130N, N156K, L161I, V250A, HA2 E179D (30%, SG 2), HA2 V193A (12%, SG 3), HA1 N156K, A195E (3%, SG 4) sowie HA1 R205K (1%, SG 5, Abb. 4). Am Ende der Influenzawelle (KW 10-13/2020) wurden nur noch 6B.1A5A-Viren nachgewiesen und am häufigsten die Varianten mit den zusätzlichen Substitutionen HA1 D187A, Q189E (n=12, SG 1), und HA1 K130N, N156K, L161I, V250A, HA2 E179D (n=11, SG 2) sowie HA2 V193A (n=2, SG 3). Zur 6B.1A5A + HA1 D187A, Q189E-Subgruppe (SG 1) gehören die Referenzviren A/Guangdong-Maonan/SWL1536/2019 und A/Hawaii/70/2019, die in der kommenden Saison 2020/21 als Impfstoffstämme eingesetzt werden.
Schwere Verläufe und Impfdurchbrüche nach A(H1N)1pdm09-Influenzavirusinfektion
Die Analyse der Altersverteilung und des Impfstatus der Patienten aus der Gruppe der schweren Verläufe zeigt, dass der Altersmedian bei den Patienten aus der SARI-Surveillance 75 Jahre (62-89 Jahre) betrug, während die Patienten mit ambulant erworbener durchschnittlich jünger (Median 29,5 Jahre, 1-61 Jahre) waren. Die Patienten mit ambulant erworbener Pneumonie waren, bis auf eine Ausnahme, nicht geimpft. Bei den übrigen Patienten mit schwerem Verlauf (Non-Sentinel) war ein älterer Patient (über 80 Jahre) mit Verdacht auf Pneumonie geimpft und ein verstorbenes Kleinkind (Altersgruppe 0-4 Jahre) war nicht geimpft. Bei den Patienten aus der SARI-Surveillance war ein älterer Patient (über 60 Jahre) mit Grunderkrankung geimpft. Bei etwa der Hälfte (8 von 23) der Virusisolate von Patienten mit schwerem Krankheitsverlauf konnte die Reaktivität gegenüber dem Impfstoffstamm A/Brisbane/02/2018 bestimmt werden und diese hatten, bis auf eine Ausnahme, eine gute Reaktivität, demzufolge eine Impfung geschützt haben könnte.
Die Analyse der HA-Gene zeigte, dass die Viren der definierten Gruppen der Impfdurchbrüche und schweren Verläufe keine separaten genetischen Cluster bilden.
A(H1N1)pdm09-Influenzaviren mit reduzierter Passgenauigkeit gegenüber dem Impfstamm
Während in der Gruppe aller analysierten Influenzaviren die 6B.1A5 + HA1 D187A, Q189E-Viren (SG 1) dominierten, wurden Viren mit reduzierter Reaktivität gegenüber dem Impfstoffstamm A/Brisbane/02/2018 (n=32) überwiegend in der 6B.1A5A + HA1 K130N, N156K, L161I, V250A, HA2 E179D-Subgruppe (n=25, SG 2) nachgewiesen. Darüber hinaus wurden in dieser Subgruppe zwei Influenzaviren mit guter Impfstammreaktivtät (mit ** markiert) detektiert, bei den übrigen Influenzaviren dieser Subgruppe (n=23) wurde die Impfstammreaktivität nicht überprüft. Weiterhin wurden Influenzaviren mit reduzierter Impfstammreaktivität in den Subgruppen 6B.1A5A + HA1 N156K, A195E (n=5, SG 4) und 6B.1A5A + HA1 D187A, Q189E (n=2, SG 1, mit * markiert) nachgewiesen. Allen Influenzaviren mit reduzierter Impfstammreaktivität ist gemeinsam, dass sie die Aminosäuresubstitution Asparagin (N) zu Lysin (K) an der Position 156 im HA1 aufweisen, welche in der Antigendomäne Sa lokalisiert ist. Im Verlauf der Saison 2019/20 wurde ein Anstieg von Viren mit der Substitution N156K und reduzierter Impfstammreaktivität nachgewiesen, was auch auf globaler Ebene beobachtet wurde.
pdm09-Viren. Quelle: © RKI") Abb. 3: Phylogenetische Analyse des Hämagglutiningens von Influenza-A(H1N1)pdm09-Viren.
Quelle: RKI
Abb. 3: Phylogenetische Analyse des Hämagglutiningens von Influenza-A(H1N1)pdm09-Viren.
Quelle: RKI
Die Referenzstämme für die einzelnen Gruppen/Subgruppen sind kursiv geschrieben, der Impfstamm (2019/20) ist zusätzlich grau unterlegt und schwarz umrahmt. Entsprechend ihres saisonalen Auftretens und ihrer Herkunft wurden die Sequenzen der Influenzaviren folgendermaßen markiert: aus dem Winter 2019/20-Non-Sentinel (schwarz), 2019/20-SARI-Surveillance (schwarz/SARI) und 2019/20-AGI-Sentinel (dunkelblau). Darüber hinaus werden in den phylogenetischen Analysen Viren von schweren akuten respiratorischen Infektionen einschließlich Todesfällen (schwere Verläufe), von Impfdurchbrüchen (ImpfD) sowie Viren mit reduzierter Impfstamm-Reaktivität (reduz. ImpfstammR) angezeigt (blau kursiv) - zum Teil auch in Kombination. Die Identifikation von Substitutionen im HA der untersuchten Influenzaviren erfolgte über Vergleiche der (abgeleiteten) Aminosäuresequenzen (A(H1N1)pdm09: H1-Nummerierung nach dem Signalpeptid beginnend). Bei den untersuchten Influenzaviren wurden aus der Saison 2019/20 die identifizierten Subgruppen- bzw. Virus-spezifischen Substitutionen im HA dargestellt, identische Sequenzen wurden vorwiegend entfernt. Substitutionen in der HA1-Untereinheit sind schwarz und in der HA2-Untereinheit blau markiert. Für die Namen der Virusisolate wurden folgende Abkürzungen verwendet: BWB: Baden-Württemberg, BAY: Bayern, BLN: Berlin, BBG: Brandenburg, BRE: Bremen, HAM: Hamburg, HES: Hessen, MVP: Mecklenburg-Vorpommern, NSA: Niedersachsen, NRW: Nordrhein-Westfalen, RPF: Rheinland-Pfalz, SAS: Sachsen, SAT: Sachsen-Anhalt, SAL: Saarland, SHO: Schleswig-Holstein, THR: Thüringen.
pdm09-Influenzaviren in der Saison 2019/20. Quelle: © RKI") Abb. 4: Anteil der HA-Varianten bei den A(H1N1)pdm09-Influenzaviren in der Saison 2019/20.
Quelle: RKI
Abb. 4: Anteil der HA-Varianten bei den A(H1N1)pdm09-Influenzaviren in der Saison 2019/20.
Quelle: RKI
Molekulare Analyse von Influenza-A(H3N2)-Viren
In die Analysen wurden folgende A(H3N2)-Influenzaviren aus der aktuellen Saison aufgenommen und charakterisiert:
In der Saison 2019/20 zirkulierten Influenza A(H3N2)-Viren zu einem großen Anteil (45% aller identifizierten Viren im AGI-Sentinel bis zur 13. KW 2020) in Deutschland. Die phylogenetische Analyse der HA-Gene zeigt, dass in der aktuellen Saison Viren der Gruppe 3C.3a dominierten, für die die Aminosäuresubstitutionen S91N, N144K, F193S, K326R im HA1 (blau umkreist) und überwiegend M17L, I149M, A201V im HA2 charakteristisch sind. Auch auf globaler Ebene zirkulierten diese Viren zu einem hohen Anteil (33% 2, 58% 3). Darüber hinaus zirkulierten in Deutschland Viren der Gruppe 3C.2a1b (HA1 E62G, K92R, H311Q), bei denen vier verschiedene Varianten (schwarz umkreist) mit zusätzlichen Aminosäureaustauschen im HA nachgewiesen wurden: 3C.2a1b + T131K (HA1 T131K, HA2 V200I), 3C.2a1b + T135K-A (HA1 T135K, T128A), 3C.2a1b + T135K-A+5 (HA1 T135K, T128A sowie A138S, G186D, D190N, F193S, S198P) und 3C.2a1b + T135K-B (HA1 T135K, T128A, S137F, A138S, F193S) (Abb. 5).
In der Gruppe aller analysierten Viren, die zu 92% aus dem AGI-Sentinel stammten, wurden überwiegend die 3C.3A-Viren (54%; 92 von 169) nachgewiesen sowie Viren der Subgruppen 3C.2a1b + T131K (27%), 3C.2a1b + T135K-A+5 mit zusätzlichen Aminosäuresubstitutionen im HA1 A138S, G186D, D190N, F193S, S198P (16%), 3C.2a1b + T135K-A (1%) sowie 3C.2a1b + T135K-B (2%, Abb. 6). Am Ende der Influenzawelle (KW 10-13/2020) wurden nur noch 3C.3A-Viren (n=22), 3C.2a1b + T131K (n=5) und 3C.2a1b + T135K-A+5 (n=4) detektiert. Viren der Subgruppe 3C.2a1b + T135K-B, zu denen auch die Referenzviren A/Hong Kong/2671/2019 und A/Hong Kong/45/2019 (Impfstoffstämme der Saison 2020/21) gehören, wurden am Ende der Saison 2019/20 in Deutschland nicht mehr detektiert, zirkulierten aber noch im März auf globaler Ebene.
Schwere Verläufe, Impfdurchbrüche und Ausbruchsuntersuchung nach Infektion mit A(H3N2)-Influenzaviren
Die Analyse der Altersverteilung und des Impfstatus der Patienten aus der Gruppe der schweren Verläufe zeigt, dass der Altersmedian bei den Patienten aus der SARI-Surveillance 77 Jahre (44-84 Jahre) betrug, während die Patienten mit ambulant erworbener durchschnittlich jünger (Median 11 Jahre, 3-60 Jahre) waren. Alle Patienten mit ambulant erworbener Pneumonie waren nicht geimpft, bei einem schwer erkrankten Patienten mit extremem Husten und zerebraler Symptomatik (Altersgruppe 35-59 Jahre, Non-Sentinel) war der Impfstatus nicht bekannt. Bei den Patienten aus der SARI-Surveillance waren sieben Patienten geimpft, alle mit einer bekannten Grunderkrankung.
Die Analyse der HA-Gene zeigt, dass die Viren der definierten Gruppen (Impfdurchbrüche und schwere Verläufe) keine separaten genetischen Cluster.
Die Analyse der HA-Gene ergab weiterhin, dass sieben der acht charakterisierten Influenzaviren von Patienten mit ambulant erworbener Pneumonie zur Gruppe der 3C.3A-Viren gehörten (Virusname blau und schwerer Verlauf). Die Analyse der Proben aus dem Ausbruch (1) zeigte, dass die Influenzaviren zur Gruppe der 3C.3A-Viren gehörten und die zusätzlich Aminosäuresubstitution K179M im HA2 aufwiesen. Darüber hinaus clustern die Influenzaviren des Ausbruchs in einem Krankenhaus mit Viren aus zwei Sentinel-Proben, welche aus nahe gelegenen Bundesländern zu einem früheren Zeitpunkt (etwa drei Wochen vorher, A/THR/3/2020, nicht gezeigt) und einem späteren Zeitpunkt (etwa sieben Wochen später, A/SAS/72/2020) entnommen wurden. Dies deutet darauf hin, dass in der Region ähnliche Viren zirkulierten. Das Influenzavirus A/SAS/72/2020, das zu einem späteren Zeitpunkt zirkulierte, zeigte zu dem einen zusätzlichen Aminosäurepolymorphismus Alanin (A)/Valin (V) an der Position 36 im HA2, welches auf eine fortschreitende Evolution dieser Viren hinweist.
A(H3N2)-Influenzaviren mit reduzierter Passgenauigkeit gegenüber dem Impfstamm
Es wurden keine A(H3N2)-Influenzaviren mit reduzierter Passgenauigkeit gegenüber dem Impfstamm detektiert.
-Viren. Quelle: © RKI") Abb. 5: Phylogenetische Analyse des Hämagglutiningens von Influenza-A(H3N2)-Viren.
Quelle: RKI
Abb. 5: Phylogenetische Analyse des Hämagglutiningens von Influenza-A(H3N2)-Viren.
Quelle: RKI
Die Referenzstämme für die einzelnen Gruppen/Subgruppen sind kursiv geschrieben, der Impfstamm (2019/20) ist zusätzlich grau unterlegt und schwarz umrahmt. Entsprechend ihres saisonalen Auftretens und ihrer Herkunft wurden die Sequenzen der Influenzaviren folgendermaßen markiert: aus dem Winter 2019/20-Non-Sentinel (schwarz), 2019/20-SARI-Surveillance (schwarz/SARI) und 2019/20-AGI-Sentinel (dunkelblau). Darüber hinaus werden in den phylogenetischen Analysen Viren von schweren akuten respiratorischen Infektionen einschließlich Todesfällen (schwere Verläufe), aus Ausbrüchen in Krankenhäusern und Altenheimen (Ausbruch) sowie von Impfdurchbrüchen (ImpfD) angezeigt (blau kursiv) - zum Teil auch in Kombination. Die Identifikation von Substitutionen im HA der untersuchten Influenzaviren erfolgte über Vergleiche der (abgeleiteten) Aminosäuresequenzen (A(H3N2): H3-Nummerierung, nach dem Signalpeptid beginnend). Bei den untersuchten Influenzaviren wurden aus der Saison 2019/20 die identifizierten Subgruppen- bzw. Virus-spezifischen Substitutionen im HA dargestellt, identische Sequenzen wurden vorwiegend entfernt. Substitutionen in der HA1-Untereinheit sind schwarz und in der HA2-Untereinheit blau markiert. Für die Namen der Virusisolate wurden folgende Abkürzungen verwendet: BWB: Baden-Württemberg, BAY: Bayern, BLN: Berlin, BBG: Brandenburg, BRE: Bremen, HAM: Hamburg, HES: Hessen, MVP: Mecklenburg-Vorpommern, NSA: Niedersachsen, NRW: Nordrhein-Westfalen, RPF: Rheinland-Pfalz, SAS: Sachsen, SAT: Sachsen-Anhalt, SAL: Saarland, SHO: Schleswig-Holstein, THR: Thüringen.
-Influenzaviren in der Saison 2019/20. Quelle: © RKI") Abb. 6: Anteil der HA-Varianten bei den A(H3N2)-Influenzaviren in der Saison 2019/20.
Quelle: RKI
Abb. 6: Anteil der HA-Varianten bei den A(H3N2)-Influenzaviren in der Saison 2019/20.
Quelle: RKI
Molekulare Analyse von Influenza-B-Viren
In die Analysen wurden folgende Influenza-B-Viren aus der aktuellen Saison aufgenommen und charakterisiert:
- Viren von Patienten mit ILI-Symptomatik (55 Patienten aus dem AGI-Sentinel und einem Patienten aus dem Non-Sentinel)
- Viren von geimpften Patienten (vier von insgesamt sieben Impfdurchbrüchen/ImpfD, darunter ein Patient mit bekannter Grunderkrankung)
- Viren mit reduzierter Reaktivität gegenüber dem Impfstoffstamm B/Colorado/06/2017 (zehn von insgesamt 23 Viren mit reduzierter Reaktivität/reduz. ImpfstammR, ≤2log2)
- Viren von drei von insgesamt vier Patienten aus der SARI-Surveillance, darunter ein Patient mit bekannter Grunderkrankung (schwerer Verlauf, siehe Tab. 1)
Während in der Saison 2019/20 Influenza-B-Viren der Victoria-Linie zu einem Anteil von 14% (aller identifizierten Viren im AGI-Sentinel bis zur 13. KW 2020) in Deutschland zirkulierten, wurde nur ein B/Yamagata-Virus (B/Berlin/5/2020, Klade 3, 3) über das AGI-Sentinel (mit geringem Virusgehalt, PCR: CT37) in einer Patientenprobe (5. KW/2020) nachgewiesen. Auch auf globaler Ebene zirkulierten vor allem B Victoria-Viren und B Yamagata-Viren (Klade 3) wurden nur vereinzelt nachgewiesen2,3.
Die phylogenetische Analyse der HA-Gene zeigt, dass in der aktuellen Saison Influenza-B/Victoria-Viren aus der Gruppe 1A(Δ162-164)B dominierten, für die die Aminosäuresubstitution K136E und die Deletion der Aminosäuren 162–164 im HA1 (schwarz umkreist) charakteristisch sind; 90% dieser Viren wiesen zusätzlich die Aminosäuresubstitution G133R (grau umkreist) auf. Darüber hinaus wurden innerhalb der 1A(Δ162-164)B-Gruppe neue Varianten identifiziert, die vier Subgruppen (grau umkreist) bilden mit den Aminosäureaustauschen HA1 G133R, E128K (SG 1), HA1 G133R, N233K (SG 2), HA1 N126K (SG 3) sowie HA1 G133R, D129N, N233S (SG 4), welche auch auf globaler Ebene beobachtet wurden. Weiterhin wurde ein 1A(Δ162-163)-Virus nachgewiesen mit Aminosäureaustauschen in den Positionen D129G, I180V im HA1, R151K im HA2 und der Deletion der Aminosäuren 162–163 im HA1 (blau umkreist, Abb. 7).
In der Gruppe aller analysierten Influenzaviren, die sich zu 92% aus dem AGI-Sentinel rekrutierten, wurden überwiegend die 1A(Δ162-164)B-Viren (98%; 61 von 62) sowie nur ein 1A(Δ162-163)-Virus nachgewiesen. Darüber hinaus wurden in der Saison 2019/20 bei den 1A(Δ162-164)B-Viren weitere Varianten mit zusätzlichen Aminosäureaustauschen identifiziert, die zu unterschiedlichen Anteilen zirkulierten: HA1 G133R (40%), HA1 G133R, E128K (22%, SG 1), HA1 G133R, N233K (21%, SG 2), HA1 N126K (8%, SG 3) sowie HA1 G133R, D129N, N233S (5%, SG 4) und ein 1A(Δ162-164)B-Virus ohne diese zusätzlichen Substitutionen (Abb. 8). Am Ende der Influenzawelle (KW 10-13/2020) wurden nur noch 1A(Δ162-164)B-Viren nachgewiesen und am häufigsten die Varianten mit den zusätzlichen Substitutionen HA1 G133R, N233K (n=6, SG 2) und HA1 G133R (n=5) sowie HA1 N126K (n=4, SG 3), G133R, E128K (n=3, SG 1) und HA1 G133R, D129N, N233S (n=2, SG 4). Zu den 1A(Δ162-164)B + HA1 G133R-Viren gehört das Referenzvirus B/Washington/02/2019, das in der kommenden Saison 2020/21 als Impfstoffstamm eingesetzt wird.
Schwere Verläufe und Impfdurchbrüche nach Infektion mit Influenza-B-Viren
Die Analyse der Altersverteilung und des Impfstatus der Patienten aus der Gruppe der schweren Verläufe zeigt, dass der Altersmedian bei den Patienten aus der SARI-Surveillance 21 Jahre (12-23 Jahre) betrug. Ein jüngerer Patient (Altersgruppe 15-34 aus der SARI-Surveillance, mit Influenza B Infektion und sonstigen Manifestationen der Atemwege, war geimpft. Im Rahmen des AGI-Sentinels wurden keine Patienten mit Influenza B Infektion und ambulant erworbener Pneumonie nachgewiesen.
Die Analyse der HA-Gene zeigt, dass die Viren der definierten Gruppen (Impfdurchbrüche, Impfstammreaktivität, schwere Verläufe) keine separaten genetischen Cluster.
Influenza-B-Viren (Victoria Linie) mit reduzierter Passgenauigkeit gegenüber dem Impfstamm
Bei den 1A(Δ162-164)B-Viren wurden bei den dominierenden Varianten, welche die zusätzlichen Aminosäureaustausche G133R, E128K sowie N233K im HA1 aufwiesen, Viren mit reduzierter Impfstammreaktivität nachgewiesen. Die Positionen 128 und 133 liegen in einer von vier Antigendomänen (120-loop, HA1 116 - 137) und der Aminosäureaustausch in der Position 233 ist in der Nähe der Rezeptorbindungsdomäne (240-loop, HA1 237 - 242) lokalisiert. Der Impfstamm der Saison 2019/20 hat diese Mutationen nicht; deswegen kann es sein, dass einzelne Viren schlechter mit dem spezifischen Antiserum reagieren, das gegen diesen Impfstamm gerichtet ist.
 Abb. 7: Phylogenetische Analyse des Hämagglutiningens von Influenza-B-Viren der Victoria-Linie.
Quelle: RKI
Abb. 7: Phylogenetische Analyse des Hämagglutiningens von Influenza-B-Viren der Victoria-Linie.
Quelle: RKI
Die Referenzstämme für die einzelnen Gruppen/Subgruppen sind kursiv geschrieben, der Impfstamm (2019/20, im trivalenten und quadrivalenten Impfstoff enthalten) ist zusätzlich grau unterlegt und schwarz umrahmt. Entsprechend ihres saisonalen Auftretens und ihrer Herkunft wurden die Sequenzen der Influenzaviren folgendermaßen markiert: aus dem Winter 2019/20-Non-Sentinel (schwarz), 2019/20-SARI-Surveillance (schwarz/SARI) und 2019/20-AGI-Sentinel (dunkelblau). Darüber hinaus werden in den phylogenetischen Analysen Viren von schweren akuten respiratorischen Infektionen einschließlich Todesfällen (schwere Verläufe), von Impfdurchbrüchen (ImpfD) sowie Viren mit reduzierter Impfstamm-Reaktivität (reduz. ImpfstammR) angezeigt (blau kursiv) - zum Teil auch in Kombination. Die Identifikation von Substitutionen im HA der untersuchten Influenzaviren erfolgte über Vergleiche der (abgeleiteten) Aminosäuresequenzen (Nummerierung nach dem Signalpeptid beginnend). Bei den untersuchten Influenzaviren wurden aus der Saison 2019/20 die identifizierten Subgruppen- bzw. Virus-spezifischen Substitutionen im HA dargestellt, identische Sequenzen wurden vorwiegend entfernt. Substitutionen in der HA1-Untereinheit sind schwarz und in der HA2-Untereinheit blau markiert. Für die Namen der Virusisolate wurden folgende Abkürzungen verwendet: BWB: Baden-Württemberg, BAY: Bayern, BLN: Berlin, BBG: Brandenburg, BRE: Bremen, HAM: Hamburg, HES: Hessen, MVP: Mecklenburg-Vorpommern, NSA: Niedersachsen, NRW: Nordrhein-Westfalen, RPF: Rheinland-Pfalz, SAS: Sachsen, SAT: Sachsen-Anhalt, SAL: Saarland, SHO: Schleswig-Holstein, THR: Thüringen.
 Abb. 8: Anteil der HA-Varianten bei den Influenza-B-Viren der Victoria-Linie in der Saison 2019/20.
Quelle: RKI
Abb. 8: Anteil der HA-Varianten bei den Influenza-B-Viren der Victoria-Linie in der Saison 2019/20.
Quelle: RKI
nach oben
Aufklärung von Ausbrüchen und Untersuchung von schweren Erkrankungsfällen
Während der Saison 2019/20 hat das NRZI an der Aufklärung von drei Influenzaausbrüchen mitgewirkt, welche sich in einem Betrieb, in einem Krankenhaus und einer Pflegeinrichtung ereigneten. Bei einem Ausbruch waren A(H3N2)-Influenzaviren (Ausbruch 1, siehe Abb. 5) ursächlich, bei den anderen Ausbrüchen konnten A(H1N1)pdm09- (Ausbruch 2) sowie A(H1N1)pdm09- und A(H3N2)-Influenzaviren (Ausbruch 3) nachgewiesen werden.
Unter den 51 Proben von schweren Erkrankungsfällen waren drei mit tödlichen Verläufen. Die Ergebnisse der PCR-Untersuchungen, Altersstruktur, Klinik, Krankheitsverlauf und vorbestehende Grunderkrankungen der Patienten (soweit bekannt) sind in Tab. 1 zusammengefasst.
Tab. 1: Im NRZ untersuchte Proben von schweren Erkrankungsfällen
 COPD: chronische obstruktive Lungenkrankheit, ITS: Intensivtherapiestation, In den Spalten 4 bis 7 wurden, soweit bekannt, alle (respiratorischen) Symptome, Befunde und weitere Angaben dargestellt, die einzeln oder in Kombination beschrieben wurden. Bei Influenza-B-Viren der Victoria-Linie wurden vier Proben von Patienten (5-14 Jahre (n=1), 15-34 Jahre (n=3)) aus der SARI-Surveillance untersucht, welche z.T. sonstige Manifestationen der Atemwege zeigten und z.T. antiviral therapiert wurden.
Quelle: RKI
COPD: chronische obstruktive Lungenkrankheit, ITS: Intensivtherapiestation, In den Spalten 4 bis 7 wurden, soweit bekannt, alle (respiratorischen) Symptome, Befunde und weitere Angaben dargestellt, die einzeln oder in Kombination beschrieben wurden. Bei Influenza-B-Viren der Victoria-Linie wurden vier Proben von Patienten (5-14 Jahre (n=1), 15-34 Jahre (n=3)) aus der SARI-Surveillance untersucht, welche z.T. sonstige Manifestationen der Atemwege zeigten und z.T. antiviral therapiert wurden.
Quelle: RKI
Untersuchungen zur antiviralen Resistenz
Im NRZI werden im Rahmen des Influenza-Sentinels entsprechend der WHO-Empfehlung kontinuierlich und zeitnah mindestens 20% der nachgewiesenen Influenza-Viren auf ihre Empfindlichkeit gegen antivirale Wirkstoffe untersucht. Dazu gehören sowohl die Ermittlung der mittleren Hemmkonzentration (IC50) der Neuraminidase-Inhibitoren Oseltamivir, Zanamivir und Peramivir mit einem fluorometrischen in house Enzym-Inhibitionstests als auch die genetische Analyse der therapeutischen Zielproteine Neuraminidase (NA), M2-Ionenkanal und Cap-abhängige Endonuklease (PA) zum Auffinden molekularer Resistenzmarker. Die erhobenen Daten fließen in die Berichterstattung des Europäischen Zentrums für die Prävention und die Kontrolle von Krankheiten (ECDC) und der WHO ein und werden zusätzlich während der Wintermonate in den Wochenberichten der AGI publiziert.
In der Saison 2019/20 wurden insgesamt 525 Influenza-A- und -B-Viren [42% A(H1N1)pdm09-, 47% A(H3N2)- und 11 Influenza B-Viren] auf ihre Resistenzeigenschaften gegenüber den NA-Inhibitoren Oseltamivir, Zanamivir und Peramivir analysiert (Tab. 2). Eine Resistenz gegen Neuraminidase-Inhibitoren, die in internationaler Übereinstimmung durch ≥10-fache (Influenza A) bzw. ≥5-fache (Influenza B) Erhöhung der IC50 von NA-Inhibitoren definiert ist, wurde in den untersuchten Viren nicht detektiert. Ein Virusisolat vom Subtyp A(H1N1)pdm09 zeigte jedoch eine 3- bzw. 7-fache Erhöhung der IC50 für Oseltamivir und Zanamivir. Hier konnte die mit verminderter Empfindlichkeit assoziierte Substitution NA-S247N detektiert werden. Diese Substitution verstärkt vor allem eine durch die Substitution NA-H275Y verursachte Resistenz gegen Oseltamivir und Peramivir und führt außerdem zu leicht verminderter Empfindlichkeit gegenüber NA-Inhibitoren.
Die Auswertung der Genomanalyse von 322 Influenza A-Viren zeigte die mit einer Resistenz gegen Adamantane (Amantadin, Rimantadin) assoziierte Substitution M2-S31N in allen untersuchten Influenza A-Viren (Tab. 2). Der in der Saison 2017 in Australien beobachtete Rückgang der Prävalenz Adamantan-resistenter Influenza A-Viren konnte damit auch in der Saison 2019/20 für die in Deutschland zirkulierenden Influenzaviren nicht bestätigt werden.
Mit dem in Deutschland noch nicht zugelassenen Wirkstoff Baloxavir marboxil steht ein Hemmer der Cap-abhängigen Endonuklease PA (PA-Inhibitor, PAI) zur Verfügung, der einen der ersten Schritte im Replikationszyklus des Influenzavirus blockiert. In klinischen Studien zeigten sich bei ca. 10% der behandelten Patienten PAI-resistente Viren, die die Substitution PA-I38T aufwiesen. Die Auswertung von 351 Genomsequenzen von Influenzaviren der Saison 2019/20 zeigte in einem A(H3N2)-Influenzavirus die Substitution PA-I38M, die ebenfalls mit einer verminderten Empfindlichkeit der Viren gegenüber diesem neuen Wirkstoff assoziiert ist (Tab. 2).
Insgesamt befindet sich die Prävalenz zirkulierender Influenzaviren mit verminderter Empfindlichkeit gegenüber NA-Inhibitoren und dem PAI Baloxavir marboxil sowohl in Deutschland als auch international auf gleichbleibend niedrigem Niveau. Derzeit gibt es keine Hinweise auf therapiebedingte Selektionierung oder therapieunabhängige Entstehung und Zirkulation von Influenzaviren, die gegen diese Wirkstoffe resistent sind. Eine natürliche Resistenz gegen die Wirkstoffklasse der Adamantane aufgrund eines Polymorphismus an Aminosäure-Position 31 des M2-Ionenkanals wurde auch in dieser Saison in allen untersuchten Influenza-A-Viren der Subtypen A(H1N1)pdm09 und A(H3N2) nachgewiesen.
Tab. 2: Resistenzsituation in Deutschland, Saison 2019-2020
 Ns: Anzahl der empfindlichen Viren, N: Anzahl der getesteten Viren, *: Substitution: PA-I38M
Quelle: RKI
Ns: Anzahl der empfindlichen Viren, N: Anzahl der getesteten Viren, *: Substitution: PA-I38M
Quelle: RKI
Zoonotische Influenzaviren
Bei einer Probe aus dem AGI Sentinel wurde ein zoonotisches Virus nachgewiesen. Ein A/sw/H1avN1 Influenzavirus von Schweinen wurde nach direkter Exposition auf ein Kind übertragen. Das Kind erholte sich nach dreitägiger Krankheit. Weitere Transmissionen fanden nicht statt4. Dies war der sechste Nachweis einer zoonotischen Übertragung von Influenzaviren der Schweine auf Menschen im NRZI. In den Jahren 2007, 2010, 2011 wurden Infektionen bei drei Patienten mit A/sw/H1avN1-Viren sowie bei einem Patienten mit A/sw/H3N2 und einem weiteren Patienten mit A/sw/H1huN2-Virus detektiert. Zwei dieser Infektionen erfolgten ebenfalls bei Kindern und eine bei einem immunsupprimierten Patienten nach Knochenmarktransplantation5. Die Kinder hatten eine ähnliche Exposition mit direktem Kontakt zu Schweinen wie bei der oben geschilderten aktuellen Transmission.
Untersuchungen zu weiteren respiratorischen Viren
Im virologischen Sentinel der AGI waren 4% (201) der 4637 Proben positiv für RSV. Im Vergleich war die Positivenrate bei den 0 bis 4 Jahre alten Patienten mit 19% (120/645) wesentlich höher. Die weiterführende genetische Charakterisierung zeigte, dass in dieser Altersgruppe 90% der Viren der RSV-Gruppe A angehörten.
HMPV wurden in 243 Sentinelproben nachgewiesen. In Proben der 0 bis 4 Jahre alten Patienten war die Positivenrate mit 12% (83/683) höher als über alle Altersgruppen (5%) und es wurden zu 69% HMPV-Gruppe A-Viren identifiziert.
Untersuchungen auf die vier humanen PIV der Typen 1 bis 4 wurden in der Saison 2020/21 neu in das Portfolio des Sentinels aufgenommen. Für humane PIV waren 189 (4%) der Sentinelproben positiv. In Bezug auf die Altersgruppe der 0 bis 4 Jahre alten Patienten war die Positivenrate mit 13% (88/677) höher als in den Proben des gesamten virologischen Sentinels. Sowohl in der Altersgruppe der 0-4 Jahre alten Patienten als auch im gesamten Sentinel wurden vorrangig PIV-1 (72%) und PIV-3 (14%-17%) in den Proben nachgewiesen.
Die virologische Surveillance wurde am 24.02.2020 um SARS-CoV-2 erweitert. Seit der 9. KW 2020 wurden insgesamt 14 SARS-CoV-2-positive Proben im Sentinel der AGI detektiert, weiterhin drei SARS-CoV-2 positive Proben im Projekt schwere akute respiratorische Infektionen. Für ausgewählte Viren wurde die Gesamtgenomsequenz erstellt und eine phylogenetischen Analyse durchgeführt, die Referenzviren aus anderen Regionen miteinbezog (Abb. 9). Gemäß der Global Initiative on Sharing All Influenza Data (GISAID) Nomenklatur6 fallen diese Viren in drei genetische Kladen: G, GR und GH. Die für das Spike Protein kodierende Genomregion ist von besonderem Interesse, da durch Impfstoffe induzierte neutralisierende Antikörper an dieses Protein binden7. In der funktionell relevanten Receptor Binding Domain (RBD, Rezeptorbindende Domäne) sind Substitutionen beschrieben, von denen zwei auch in den hier dargestellten deutschen Isolaten auftraten (Abb. 9).
Mit den Maßnahmen zur Reduzierung der weiteren SARS-Cov-2-Ausbreitung im Frühjahr gingen die Nachweise respiratorischer Viren drastisch zurück. In KW18/2020 und KW22/2020 wurde kein respiratorisches Virus nachgewiesen, in den dazwischenliegenden Wochen nur vereinzelt RSV und Rhinoviren und in KW24/2020 und KW25/2020 vereinzelt RSV und PIV. Nach der Lockerung der Maßnahmen nahm die Zirkulation der Rhinoviren ab KW23/2020 zu. Mit Ausnahme von drei Influenzavirusnachweisen (ein zoonotisches A/sw/H1avN1 Influenzavirus im Juni und zwei saisonale Influenzaviren im September: A(H3N2) und B/Victoria Linie) wurden in der gesamten Zeit seit KW26/2020 keine anderen der oben erwähnten respiratorischen Viren nachgewiesen (kein RSV, kein PIV und kein HMPV).
. Quelle: © RKI") Abb. 9: Phylogenetische Analyse von SARS-CoV-2, die in Sentinelproben der AGI nachgewiesen wurden (schwarz umrahmt). Der betrachtete Genomabschnitt umfasst nahezu die volle SARS-CoV-2 Genomlänge (nt266 – nt29674). Referenzsequenzen (kursiv) wurden in die Analyse miteinbezogen und sind farblich gemäß ihrer GISAID- Kladen-Zugehörigkeit gekennzeichnet.
Quelle: RKI
Abb. 9: Phylogenetische Analyse von SARS-CoV-2, die in Sentinelproben der AGI nachgewiesen wurden (schwarz umrahmt). Der betrachtete Genomabschnitt umfasst nahezu die volle SARS-CoV-2 Genomlänge (nt266 – nt29674). Referenzsequenzen (kursiv) wurden in die Analyse miteinbezogen und sind farblich gemäß ihrer GISAID- Kladen-Zugehörigkeit gekennzeichnet.
Quelle: RKI
nach oben
Zusammenfassung
Die Grippewelle der Saison 2019/20 begann in der 2. KW 2020, erreichte in der 5. bis 7. KW 2020 ihren Höhepunkt und endete nach Definition der Arbeitsgemeinschaft Influenza in der 12. KW 2020. Mit elf Wochen war die Grippewelle der Saison 2019/20 im Vergleich zu den vergangenen fünf Saisons (11-15 Wochen) kurz, was im Zusammenhang mit den bundesweiten Maßnahmen zur Eindämmung der COVID-19-Pandemie zu stehen scheint. Insgesamt verlief die Grippewelle moderat. Es zirkulierten hauptsächlich die beiden Influenza-A-Subtypen; jedoch wurden im Unterschied zu der Vorsaison auch Influenza-B-Viren der Victoria-Linie detektiert. Bei einem Drittel der A(H1N1)pdm09 Influenzaviren war die Passfähigkeit der Impfstämme reduziert, auch bei wenigen Influenza-B-Viren der Victoria Linie, dennoch waren die Impfstoffe gegen alle zirkulierenden Viren wirksam und die Wirksamkeit wurde höher als in den Vorsaisons geschätzt. Es wurden keine Resistenzen gegen Neuraminidaseinhibitoren festgestellt. Ein Virus hatte die Substitution PA-I38M, die mit einer verminderten Empfindlichkeit der Viren gegenüber Baloxavir marboxil assoziiert ist. Alle untersuchten Viren waren resistent gegen Adamantane.
Danksagung
Wir bedanken uns bei allen Sentinelpraxen, dem Helios Klinikum Emil von Behring, Berlin, und dem Diakonie Krankenhaus, Bad Kreuznach für die Einsendung von Proben. Wir danken folgenden Instituten für die Einsendung von Influenzavirusisolaten: Institut für Virologie, Universitätsklinikum Ulm, Institut für Mikrobiologie und Hygiene, Universität Freiburg, Labor Enders, Stuttgart, Landesuntersuchungsanstalt für das Gesundheits- und Veterinärwesen Sachsen, Dresden, Landesgesundheitsamt Baden-Württemberg, Stuttgart, Landesamt für Verbraucherschutz Sachsen-Anhalt, Magdeburg, Niedersächsisches Landesgesundheitsamt, Hannover, Thüringer Landesamt für Verbraucherschutz, Bad Langensalza. Wir danken den Autoren von GISAID für die Sequenzen, die für vergleichende Analysen genutzt wurden. Weiterhin bedanken wir uns bei Mareen Adam, Heike Fischer, Susi Hafemann, Ute Hopf-Guevara, Carmen Karstädt-Schulze, Katja-Irena Madaj, Jeanette Milde, Bettina Mischke, Christine Spingies, Anneliese Schindel, Kathrin Seidel, Nathalie Tollard, Robert Koch Institute, Berlin, Germany für Assistenz bei den Laboruntersuchungen. Frau Ute Preuß, Abt. 3 Infektionsepidemiologie, FG 36 Respiratorisch übertragbare Erkrankungen, Robert Koch-Institut danken wir für die Erstellung der Abb. 1.
nach oben
Literatur
1Buda S, Preuß U, Wedde M, Dürrwald R: Wirksamkeit der saisonalen Influenzaimpfung bei ambulant behandelten Patienten in der Saison 2019/20 in Deutschland. Epid Bull 2020; 45:3-6 | DOI 10.25646/7159
2McCauley J, Daniels R, Lin YP, Harvey R, Galiano M, Xiang Z, Whittaker L, Halai C, Cross K, Rattigan A, Ermetal B, Lilley A, Dai M, Bennett M, Hussain S. Report prepared for the WHO annual consultation on the composition of influenza vaccine for the Northern Hemisphere 2020-2021. 24th-27th February 2020 (https://www.crick.ac.uk/sites/default/files/2020-03/Crick%20WIC%20Report%20Feb%202020.pdf)
3McCauley J, Daniels R, Lin YP, Harvey R, Galiano M, Xiang Z, Ermetal B, Rattigan A, Cross K, Whittaker L, Lilley A, Dai M, Bennett M, Halai C, Hussain S. Report prepared for the WHO annual consultation on the composition of influenza vaccine for the Southern Hemisphere 2021. 24th-27th February 2020 (https://www.crick.ac.uk/sites/default/files/2020-10/Crick_SH2021%20report_Full%20seasonal.pdf)
4Dürrwald R, Wedde M, Biere B, Oh D-Y, Heßler-Klee M, Geidel C, Volmer R, Hauri AM, Gerst K, Thürmer A, Appelt S, Reiche J, Duwe S, Buda S, Wolff T, Haas W. Zoonotic infection with swine A/H1avN1 influenza virus in a child, Germany, June 2020. Euro Surveill. 2020;25(42):pii=2001638
5Buda S, Haas W, Baillot A, Beyrer K, Monazahian M, Pulz M, Benzler J, Harder T, Schweiger B (2011) Humane Fälle mit Infektion durch Schweineinfluenzaviren. Epid Bull 2011,39:357-359.
6Alm, E., Broberg, E.K., Connor, T., Hodcroft, E.B., Komissarov, A.B., Maurer-Stroh, S., Melidou, A., Neher, R.A., O'Toole, A., Pereyaslov, D., The WHO European Region sequencing laboratories and GISAID EpiCoV group1 (2020). Geographical and temporal distribution of SARS-CoV-2 clades in the WHO European Region, January to June 2020. Euro Surveill. 2020;25(32):pii=2001410.
7Liu, L., Wang, P., Nair, M.S., Yu, J., Rapp, M., Wang, Q., Luo, Y., Chan, J.F., Sahi, V., Figueroa, A., et al. (2020). Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature 584, 450-456.
nach oben
Autorinnen und Autoren
Marianne Wedde, Susanne Duwe, Barbara Biere, Sophie Köndgen, Janine Reiche, Djin-Ye Oh, Ralf Dürrwald
Robert Koch-Institut, Abt. 1 Infektionskrankheiten, FG 17 Influenzaviren und weitere Viren des Respirationstraktes
Korrespondenz: duerrwaldr@rki.de
nach oben
Interessenkonflikt
Die Autorinnen und Autoren erklären, dass kein Interessenkonflikt besteht.
nach oben


 Reife Virionen (rote Hülle) sammeln sich an der Oberfläche eines T-Lymphozyten(Wirtszelle). Transmissions-Elektronenmikroskopie, Ultradünnschnitt. Quelle: © H.R. Gelderblom/RKI")
 auf Blutagar ohne Hämolyse. Quelle: © Bredow/RKI")









